 "
"
Team:BV CAPS Kansas/Solutions
From 2013hs.igem.org
(Difference between revisions)
Meaton2222 (Talk | contribs) |
Meaton2222 (Talk | contribs) |
||
| Line 1,179: | Line 1,179: | ||
By overexpressing the catalytic protein pyruvate kinase, we hope to increase pyruvate within the cell. This should allow for an increase in activity within the alkane fatty-acid pathway because of pyruvates conversion to acetyl-CoA. See below for the pathway. <br><br><br> | By overexpressing the catalytic protein pyruvate kinase, we hope to increase pyruvate within the cell. This should allow for an increase in activity within the alkane fatty-acid pathway because of pyruvates conversion to acetyl-CoA. See below for the pathway. <br><br><br> | ||
| - | <center><a ><img src="https://static.igem.org/mediawiki/2013hs/ | + | <center><a ><img src="https://static.igem.org/mediawiki/2013hs/f/f5/Metabolic_Pathway_New.png" width="400"></a></center><br><br> |
To increase the efficiency of the production of alkanes, we chose to use cyanobacteria for our chassis. This autotroph doesn’t require an expensive carbon fuel source; instead, it uses one of the world’s harmful surpluses – carbon dioxide. We are using <i>Synechocystis PCC 6803</i> and <i>Synechococcus PCC 6801</i>. <i>S. 6803</i> was the first photosynthetic organism to have its genome sequenced. As a model organism, it contains one chromosome, seven plasmids, and is both phototrophic and heterotrophic. <br><br><br> | To increase the efficiency of the production of alkanes, we chose to use cyanobacteria for our chassis. This autotroph doesn’t require an expensive carbon fuel source; instead, it uses one of the world’s harmful surpluses – carbon dioxide. We are using <i>Synechocystis PCC 6803</i> and <i>Synechococcus PCC 6801</i>. <i>S. 6803</i> was the first photosynthetic organism to have its genome sequenced. As a model organism, it contains one chromosome, seven plasmids, and is both phototrophic and heterotrophic. <br><br><br> | ||
Revision as of 17:25, 21 June 2013









BV CAPS iGEM Tweets
Thanks!





Solutions
Attempted solutions
Renewable energy is a growing industry. This industry is anticipated to supplement and, ultimately, replace non-renewable fuels. The United States Energy Independence and Security Act of 2007 proclaimed a goal to produce 36 billion gallons of renewable fuel, with 16 billion being cellulosic ethanol, by the year 2022. This was the first major step in the advancement of biofuels.
Biofuels are energy produced from renewable resources that have a balanced carbon dioxide emission-absorption trade-off. The optimal materials to make biofuels are plant biomass, vegetable oil, and especially municipal and industrial wastes. The major classifications of biofuel are biodiesel, biogas, and bioethanol. Biodiesel is made by the emulsification of vegetable oils and other fats. Commonly, biodiesel suplements petroleum-based fuel. The second class, biogas, is the decomposition of organic matter. This creates a mixture of methane and carbon dioxide that is used to generate steam and electricity. Finally, there is bioethanol. This is made by fermenting the sugars in corn or other agricultural residues. Bioethanol can be used in internal-combustion engines.
Currently, the most common biofuel is corn-based ethanol. This first-generation biofuel accounted for 2.7% of transportation fuel in 2010. Ethanol, however, is quite expensive to produce and not nearly as efficient as gasoline or diesel. Another concern is the use of land to produce corn for fuel, rather than food.
Second generation biofuels, those derived from lignocellulosic biomass (dry plant matter), are the most abundant carbon fuel source. Unfortunately, they, like first generation fuels, require land, energy, and nutrient investment. A major barrier to production is the high energy required to extract trapped sugars during fermentation. Even then, there is a high percentage of pentoses (five carbon monosaccharides) that are even more difficult to ferment. A low cost, low input way to create renewable biofuel is still being sought after.
Third-generation biofuels are a source of hope. Many of these utilize photosynthetic microbes, like cyanobacteria. This method requires only sunlight and adequate growing condition. The microbes do not need much space or nutrient input. However, biofuel output through this method is relatively low. The possibilities here sparked our interest.
Our Solutions
Most of the energy in both diesel and gasoline comes from alkanes. A cell makes alkanes by the fatty-acid pathway. A main resource needed for the fatty-acid pathway, acetyl-CoA, is produced by glycolysis. We hypothesize that if we increase the resources required for the production of alkanes, then the cell will produce more alkanes.
By overexpressing the catalytic protein pyruvate kinase, we hope to increase pyruvate within the cell. This should allow for an increase in activity within the alkane fatty-acid pathway because of pyruvates conversion to acetyl-CoA. See below for the pathway.
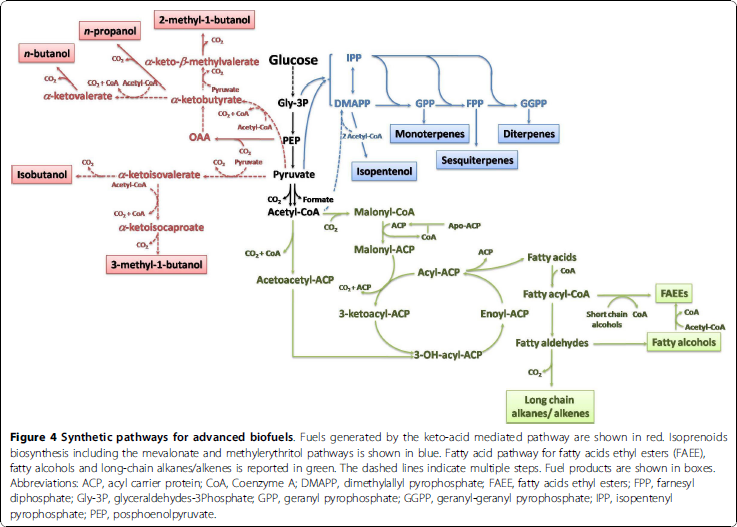
To increase the efficiency of the production of alkanes, we chose to use cyanobacteria for our chassis. This autotroph doesn’t require an expensive carbon fuel source; instead, it uses one of the world’s harmful surpluses – carbon dioxide. We are using Synechocystis PCC 6803 and Synechococcus PCC 6801. S. 6803 was the first photosynthetic organism to have its genome sequenced. As a model organism, it contains one chromosome, seven plasmids, and is both phototrophic and heterotrophic.

Therefore, we plan to increase the production of alkanes by reducing the bottleneck of limited resources. By placing an overactive system in cyanobacteria, we will create more resources to create alkanes. We’ll used a broad-host range plasmid from Hawaii’s 2008 iGEM team (BBa_K125000) to be the back bone for our two highly active pyruvate genes. One is derived from rabbit muscle (BBa_K977000) and the other is a yeast mutant (BBa_K977001).
The Alkane Pathway Explained Step by Step:
- Hexokinase, alongside ATP, phosphorylates (adds a phosphate group to) a molecule of glucose, yielding ADP Glucose 6-Phosphate
- Phosphoglucose Isomerase isomerizes (changes form of) Glucose 6-Phosphate into Fructose 6-Phosphate
- Phosphofructokinase, alongside ATP, phosphorylates Fructose 6-Phosphate, yielding ADP and Fructose 1, 6-Biphosphate
- Fructose Biphosphate Aldolase cleaves Fructose 1, 6-Biphosphate into Glyceraldehyde 3-Phosphate and Dihydroxyacetone Phosphate
- Triosephosphate Isomerase isomerizes Dihydroxyacetone Phosphate into another Glyceraldehyde 3-Phosphate
- Glyceraldehyde Phosphate Dehydrogenase, alondside NAD+ and H2(PO4), dehydrogenizes (removes hydrogen from) Glyceraldehyde 3-Phosphate, yielding NADH/H+ and 1, 3-Biphosphoglycerate
- Phosphoglycerate Kinase, alongside ADP, dephosphorylates 1, 3-Biphosphoglycerate, yielding ATP and 3-Phosphoglycerate
- Phosphoglycerate Mutase converts 3-Phosphoglycerate into 2-Phosphoglycerate
- Enolase pulls a molecule of H2O from 2-Phosphoglycerate, yielding Phosphoenolpyruvate (PEP)
- Pyruvate Kinase, alongside ADP and H+, dephosphorylates PEP, yielding ATP and Pyruvate
- Pyruvate Dehydrogenase, alongside NAD+, dehydrogenizes Pyruvate, yielding NADH/H+ and Acetyl Coenzyme-A
- Acetyl CoA Carboxylase, alongside ATP and a carboxyl group (H(CO3)), carboxylizes Acetyl CoA, yielding Malonyl Coenzyme-A
- Beta-Ketoacyl-ACP Synthase and Acetyl CoA react with Malonyl CoA, yielding CO2 and Acetoacetyl-ACP
- 2NADPH and Malonyl-ACP react with Acetoacetyl-ACP, facilitating hydrocarbon chain elongation, yielding CO2, NADP+, and Acyl-ACP
- Acyl-ACP Reductase, alongside NADPH, reduces (changes the oxidation state of) Acyl-ACP, yielding NADP+ and Acyl-AH
- Aldehyde Decarbonylase decarbonylizes (removes a carbonyl group (CO) from) Acyl-AH, yielding an Alkane